Abstract
In Senegal, despite numerous malaria control interventions, transmission is still seasonal. Malaria transmission depends on the presence of infectious parasites in the sexual stage in human peripheral blood. Immune responses acquired naturally at these or other stages can affect malaria transmission, resulting in protection against malaria, reduced transmission, and also form the basis for the development of transmission-blocking vaccines. To evaluate the antibody response profile against the asexual antigens PfAMA1, PfMSP119, Pf GLURP R2 and the sexual antigens Pfs230C1, Pfs48.45.6C in inhabitants naturally exposed to malaria in areas with different levels of transmission in Senegal. A cross-sectional study was carried out at the end of the transmission season in central (Keur Socé) and southern (Saraya) Senegal in 2018. We included 1106 asymptomatic volunteers aged 5 and over. Capillary blood was collected from each participant for an RDT, 2 slides for microscopy and a dried blood spot samples for immunology. A Luminex serological multiplex bead assay was then used to assess Plasmodium falciparum seroprevalence Our study population was characterized by a very young population with a median age of 12 15 years. The parasite prevalence of Plasmodium falciparum was 21.75% and 2.75% by RDT and 22.1% and 2.2% by microscopy for the southern and central regions respectively. Two other plasmodial species were found in Saraya, with prevalences of 1.61% for P. malariae and 0.18% for P. ovale. The mean seroprevalences of antibodies against three asexual blood-stage antigens (PfAMA1, PfGLURP and PfMSP119) and two sexualstage antigens (Pfs48.45.6C and Pfs230C1) were significantly higher in Saraya. In Keur Socé, the mean seroprevalence of antibodies against the PfAMA1 antigen was highest (1.83%), while in Saraya, PfMSP119 was highest (49.91%). The antigenicity of these proteins depended on endemicity levels, as antibody prevalence was statistically different in the two sites and increased with transmission intensity. With the exception of anti-Pfs48.45.6C antibody levels, all other antibody responses increased with age. Overall, these data indicate that the seroprevalence and antivody levels of individuals with antibodies recognizing all five antigens increase with exposure to infection, and that these antibodies may contribute to immunity against parasites. Children receiving SMC should also be monitored, as we have noted a loss of immunity in this group.
Keywords
Antigens, Antibodies, Immunology, Plasmodium Falciparum, Transmission
1. Introduction
Malaria continues to represent a major burden for global health, with 409,000 malaria-related deaths reported in 2019 (WHO, 2020), causing substantial morbidity and mortality, particularly in sub-Saharan Africa
| [1] | Combemorel P. Le cycle de vie de Plasmodium falciparum | Planet-Vie [The life cycle of Plasmodium falciparum | Planet-Life]. 2019 [cité 30 nov 2025]. |
[1]
.
Plasmodium falciparum is responsible for the majority of severe malaria cases. The life cycle of
P. falciparum includes asexual stages, which are responsible for clinical symptoms, and sexual stages (gametocytes), which are essential for transmission of the parasite through mosquito vectors
| [2] | Barry A, Hansen D. Naturally acquired immunity to malaria. Parasitology. févr 2016; 143(2): 125-8. |
[2]
.
Malaria transmission depends on the presence of infectious gametocytes in human peripheral blood
| [3] | Epidemiology and Infectivity of Plasmodium falciparum and Plasmodium vivax Gametocytes in Relation to Malaria Control and Elimination | Clinical Microbiology Reviews [Internet]. [cité 30 nov 2025]. |
[3]
. Naturally acquired immunity against both the sexual and asexual stages of the parasite can modulate this transmission, notably by reducing the density of asexual parasites that produce gametocytes
| [4] | Khan SM, Franke-Fayard B, Mair GR, Lasonder E, Janse CJ, Mann M, et al. Proteome analysis of separated male and female gametocytes reveals novel sex-specific Plasmodium biology. Cell. 3 juin 2005; 121(5): 675-87. |
[4]
. During their maturation, gametocytes undergo substantial changes in protein expression, including the upregulation of antigens essential for parasite development in the mosquito
| [5] | van Dijk MR, Janse CJ, Thompson J, Waters AP, Braks JA, Dodemont HJ, et al. A central role for P48/45 in malaria parasite male gamete fertility. Cell. 12 janv 2001; 104(1): 153-64. |
[5]
, as well as specific surface proteins involved in fertilization and sporogonic stages
| [6] | Tachibana M, Miura K, Takashima E, Morita M, Nagaoka H, Zhou L, et al. Identification of domains within Pfs230 that elicit transmission blocking antibody responses. Vaccine. 22 mars 2019; 37(13): 1799-806. |
[6]
. A proportion of gametocytes die before being transmitted, exposing their antigens to the host immune system and inducing the production of specific antibodies, some of which can block fertilization or inhibit parasite development within the mosquito
| [7] | Sauerwein RW, Bousema T. Transmission blocking malaria vaccines: Assays and candidates in clinical development. Vaccine. 22 déc 2015; 33(52): 7476-82. |
| [8] | Yazdani SS, Mukherjee P, Chauhan VS, Chitnis CE. Immune responses to asexual blood-stages of malaria parasites. Curr Mol Med. mars 2006; 6(2): 187-203. |
[7, 8]
. Thus, naturally acquired immunity targeting sexual stages may limit malaria transmission and provides a crucial foundation for the development of transmission-blocking vaccines
| [3] | Epidemiology and Infectivity of Plasmodium falciparum and Plasmodium vivax Gametocytes in Relation to Malaria Control and Elimination | Clinical Microbiology Reviews [Internet]. [cité 30 nov 2025]. |
| [8] | Yazdani SS, Mukherjee P, Chauhan VS, Chitnis CE. Immune responses to asexual blood-stages of malaria parasites. Curr Mol Med. mars 2006; 6(2): 187-203. |
[3, 8]
.
In populations living in endemic areas, repeated exposure to the parasite allows the development of so-called “naturally acquired” immunity. This immunity does not lead to complete parasite clearance, but it helps limit parasite burden, reduce the frequency and severity of clinical episodes, and potentially influence transmission
| [9] | Healer J, Chiu CY, Hansen DS. Mechanisms of naturally acquired immunity to P. falciparum and approaches to identify merozoite antigen targets. Parasitology. juin 2018; 145(7): 839-47. |
| [10] | De Jong RM, Tebeje SK, Meerstein-Kessel L, Tadesse FG, Jore MM, Stone W, et al. Immunity against sexual stage Plasmodium falciparum and Plasmodium vivax parasites. Immunol Rev. janv 2020; 293(1): 190-215. |
[9, 10]
.
Historically, most studies have focused on immunity directed against the asexual blood stages of the parasite, as these are the stages that cause disease
| [10] | De Jong RM, Tebeje SK, Meerstein-Kessel L, Tadesse FG, Jore MM, Stone W, et al. Immunity against sexual stage Plasmodium falciparum and Plasmodium vivax parasites. Immunol Rev. janv 2020; 293(1): 190-215. |
[10]
. However, recent work has shown that immune response particularly humoral responses can also target antigens of the sexual stages (gametocytes), which could reduce not only disease in the human host but also transmission to the mosquito
| [11] | Ouédraogo AL, Roeffen W, Luty AJF, de Vlas SJ, Nebie I, Ilboudo-Sanogo E, et al. Naturally Acquired Immune Responses to Plasmodium falciparum Sexual Stage Antigens Pfs48/45 and Pfs230 in an Area of Seasonal Transmission ▿. Infect Immun. déc 2011; 79(12): 4957-64. |
| [12] | Kengne-Ouafo JA, Sutherland CJ, Binka FN, Awandare GA, Urban BC, Dinko B. Immune Responses to the Sexual Stages of Plasmodium falciparum Parasites. Front Immunol [Internet]. 1 févr 2019 [cité 30 nov 2025]; 10. |
[11, 12]
.
Thus, studying natural immunity against sexual and asexual stage antigens is of dual interest: on the one hand, to understand the mechanisms of premunition and immune adaptation in exposed populations; on the other hand, to identify potential targets for vaccine strategie particularly (transmission-blocking vaccines)
| [12] | Kengne-Ouafo JA, Sutherland CJ, Binka FN, Awandare GA, Urban BC, Dinko B. Immune Responses to the Sexual Stages of Plasmodium falciparum Parasites. Front Immunol [Internet]. 1 févr 2019 [cité 30 nov 2025]; 10. |
| [13] | Linn AM, Ndiaye Y, Hennessee I, Gaye S, Linn P, Nordstrom K, et al. Reduction in symptomatic malaria prevalence through proactive community treatment in rural Senegal. Trop Med Int Health TM IH. nov 2015; 20(11): 1438-46. |
[12, 13]
.
In this context, rural areas of Senegal, such as the southern region (Saraya) and the central region (Keur Soce), constitute particularly relevant study sites: the heterogeneity of transmission intensity, the diversity of exposure profiles, and the history of repeated infections provide a unique opportunity to assess the acquisition, quality, and dynamics of naturally acquired immune responses to the various stages of P. falciparum.
The present work therefore aims to characterize, in these two geographically and epidemiologically distinct areas, the magnitude and nature of naturally acquired immune responses directed against PfAMA1, PfMSP1, GLURP, Pfs230C1, and Pfs48.45.6C antigens—representative of the asexual and sexual stages of P. falciparum—in order to better understand their potential role in protection against disease and transmission.”
2. Materials and Methods
2.1. Study
Study design: We conducted two cross-sectional surveys in January 2018 in two health districts (Saraya and Ndoffane), characterized by high and low malaria transmission, respectively. The District of Saraya, located in the extreme southeast of Senegal in the Kédougou region, is a very remote, rural area. With a surface area of 6800 km2, it borders Mali and Guinea. During the 2013 census conducted in the district, the population was estimated at 52,590 permanent residents, not including the influx of small-scale artisanal miners into the area
| [14] | Ndiaye Y, Ndiaye JLA, Cisse B, Blanas D, Bassene J, Manga IA, et al. Community case management in malaria: review and perspectives after four years of operational experience in Saraya district, south-east Senegal. Malar J. 12 juill 2013; 12: 240. |
[14]
. The population density is less than 9 people per km2 and 70% of this population lives more than 5 km from a health post or center
| [15] | Agence Nationale de la Statistique et de la Démographie (ANSD) et ICF International. Enquête Démographique et de Santé à Indicateurs Multiples Sénégal (EDS-MICS) 2010-2011 [National Agency for Statistics and Demography (ANSD) and ICF International. Senegal Multiple Indicator Cluster Survey (MICS-EDS) 2010-2011]. Calverton, Maryland, USA; 2012. (ANSD et ICF International). |
[15]
. Infant mortality was estimated at 154 deaths per 1,000 live births
| [14] | Ndiaye Y, Ndiaye JLA, Cisse B, Blanas D, Bassene J, Manga IA, et al. Community case management in malaria: review and perspectives after four years of operational experience in Saraya district, south-east Senegal. Malar J. 12 juill 2013; 12: 240. |
[14]
. It has been noted that Kedougou has the highest prevalence of
P. falciparum in children under 5 years of age in Senegal (13.5% compared with the national average of 3%)
| [16] | Ndiath M, Faye B, Cisse B, Ndiaye JL, Gomis JF, Dia AT, et al. Identifying malaria hotspots in Keur Soce health and demographic surveillance site in context of low transmission. Malar J. 24 nov 2014; 13(1): 453. |
[16]
. The community of Keur Socé is located in a rural area of the Kaolack region, in the Ndoffane health district. It lies 230 km from Dakar in the Sudano-Sahelian region of Senegal. The site's ecology is characterized by an alternating long dry season from November to June and a rainy season from July to October. In August 2013, the population was 32,601 with 2,371 households
| [17] | Sylla K, Tine RCK, Ndiaye M, Sow D, Sarr A, Mbuyi MLT, et al. Sero-epidemiological evaluation of Plasmodium falciparum malaria in Senegal. Malar J. 16 juill 2015; 14(1): 275. |
[17]
.
2.2. Study Population
A cross-sectional survey was conducted in the localities of Saraya and Keur Socé. Villages were selected based on the registers of reported cases within each health district, and a random draw was performed to identify the households to be included. In Keur Socé, the entire community was involved due to the low malaria incidence. In contrast, in Saraya, where malaria cases were more numerous, eight villages were selected: Diakha Madina, Saroudia, Badioula, Diakha Guenedji, Missirah Dantila, Nafadji, Moussala, and Diakhaba.
Children aged 5 years and above, as well as adults residing in the selected households who provided informed consent, were included in the study, regardless of whether they were positive or negative for malaria parasites according to the rapid diagnostic test. Any individual presenting an illness other than malaria was excluded from the study.
2.3. Sample Size Determination
The sample size was calculated on the basis of the seroprevalence of anti-CSP antibodies of 38% reported in Senegal in 2015
| [17] | Sylla K, Tine RCK, Ndiaye M, Sow D, Sarr A, Mbuyi MLT, et al. Sero-epidemiological evaluation of Plasmodium falciparum malaria in Senegal. Malar J. 16 juill 2015; 14(1): 275. |
[17]
, with a margin error of 5% and a power of 80% to give a minimum sample size of 1259 participants needed in the two study sites, using the following formula:
n
2.4. Data Collection
An informed consent questionnaire was administered to collect individual sociodemographic information (age, sex, axillary temperature, ownership of an insecticide-treated net, and use of the net the previous night). Capillary blood samples were then collected from eligible participants for the detection of Plasmodium spp. using a rapid diagnostic test (RDT), the preparation of two thick blood smears and one thin smear for microscopic examination, as well as the deposition of a few drops of blood onto filter paper. The filter papers were air-dried and individually placed in plastic bags containing silica gel. Samples were stored at +4°C until further use for Luminex serological analyses. Anemia status was also assessed using the HemoCue device. Finally, all samples were transported to the laboratory in accordance with standard preservation procedures.
2.5. Laboratory Analysis
2.5.1. Parasite Detection
Plasmodium species identification and parasitaemia were determined using 100X oil immersion microscopy by two experienced microscopists. Thin and thick blood smears were processed for Giemsa staining and evaluated using a WHO protocol
| [18] | World Health Organization. World malaria report 2020 [Internet]. [cité 17 oct 2025]. Report No.: 978-92-4-001579-1. |
[18]
.
Plasmodium species were identified after evaluation of the thin layers, and parasite density was estimated using the thick layers. Parasite density was determined as the percentage of infected erythrocytes counted per 200 white blood cells (WBC) based on a WHO protocol
| [18] | World Health Organization. World malaria report 2020 [Internet]. [cité 17 oct 2025]. Report No.: 978-92-4-001579-1. |
[18]
, and converted to parasites/μL assuming a density of 8,000 white blood cells/μL of blood. The average parasite density of the two readings was recorded, and a third microscopist consulted in the event of disagreement.
2.5.2. Antibody Response to PfMSP119, PfAMA1, PfGLURP and Pfs230 and Pfs48.45.6C parasite Sexual Stage ANTIGENS
The multiplex bead-based Luminex serological assay enabled us to quantify antibodies directed against our antigens PfMSP1
19, PfAMA1, PfGLURP, Pfs230, and Pfs48.45.6C (
Table 1 [19-21]).
Table 1. Antigen Information.
Antigen | Full Name | Parasite Stage | Reference / Notes |
PfMSP1 19 | Merozoite Surface Protein 1, 19 kDa C terminal fragment | Asexual blood stage (merozoite surface) | MSP1 19 remains anchored on invading merozoites after processing; a major vaccine candidate | [19] | Muerhoff AS, Birkenmeyer LG, Coffey R, Dille BJ, Barnwell JW, Collins WE, et al. Detection of Plasmodium falciparum, P. vivax, P. ovale, and P. malariae Merozoite Surface Protein 1-p19 Antibodies in Human Malaria Patients and Experimentally Infected Nonhuman Primates. Clin Vaccine Immunol CVI. oct 2010; 17(10): 1631-8. |
[19] |
PfAMA1 | Apical Membrane Antigen 1 | Asexual blood stage (merozoite apical organelles) | Commonly used in antibody serology panels | [20] | Rogier E, van den Hoogen L, Herman C, Gurrala K, Joseph V, Stresman G, et al. High-throughput malaria serosurveillance using a one-step multiplex bead assay. Malar J. 4 déc 2019; 18(1): 402. |
[20] |
PfGLURP | Glutamate Rich Protein (R0 region, blood stage) | Asexual blood stage | Often combined with asexual antigens in sero prevalence studies | [20] | Rogier E, van den Hoogen L, Herman C, Gurrala K, Joseph V, Stresman G, et al. High-throughput malaria serosurveillance using a one-step multiplex bead assay. Malar J. 4 déc 2019; 18(1): 402. |
[20] |
Pfs230 | Pfs230 (gametocyte/gamete membrane protein) | Sexual stage (gametocytes/gametes) | A sexual stage antigen targeted by transmission blocking antibodies | [21] | Eacret JS, Parzych EM, Gonzales DM, Burns JM. Inclusion of an optimized Plasmodium falciparum merozoite surface protein 2-based antigen in a trivalent, multi-stage malaria vaccine. J Immunol Baltim Md 1950. 15 avr 2021; 206(8): 1817-31. |
[21] |
Pfs48.45.6C | Pfs48.45, C terminal 6 Cys domain fragment | Sexual stage (gametocytes/gametes) | Potent transmission blocking domain, target of mAb 85RF45.1 |
The 230-CMB antigen was produced by Fraunhofer; it contains domain 1 and part of the pro-domain of Pfs230 and differs slightly from Pfs230C1 in the amino-acid region covered. The Pfs48/45-6C antigen is produced in insect cells (Radboud UMC, Nijmegen, the Netherlands). (see supplementary document for the Luminex protocol).
2.6. Statistical Analysis
Data were processed using Microsoft Excel 2010, with patients' first and last names coded. Study participants were classified into age groups (5-10 years, 11-25 years, 26-40 years and over 40 years). All statistical analyses were performed using R and STATA software. Quantitative variables were presented as mean ± standard deviation (SD) or median ± interquartile range (IQR), depending on the normality of the variables under consideration, while qualitative variables were presented as headcounts and percentages. As the antibody data did not meet the parametric test assumptions, we used non-parametric tests: Wilcoxon test (comparison of two medians) and Kruskal-Wallis test (comparison of three or more medians). The Chi squared test was used to compare percentages. Spearman's test was used to analyze correlation between variables. The significance threshold was set at 0.05.
The positivity threshold was calculated based on the negative population. We used the average of 21 negative samples plus 3 times the standard deviation. This gives a cut-off guaranteeing very high specificity (~99.7%) in a normal distribution. Negative controls consist of malaria-naïve European individuals residing in the United Kingdom who have never travelled to malaria-endemic regions.
3. Results
3.1. Socio-demographic Characteristics
The main characteristics of the study participants are presented in
Table 2. A total of 1,106 individuals from Keur Socé and Saraya were included. No significant differences were observed between the two localities regarding age (P = 0.45) and mean hemoglobin concentration (Hb) (P = 0.20). However, the sex distribution differed significantly, with a higher proportion of males in Saraya and the opposite pattern in Keur Socé (P < 0.001). Similarly, the proportion of febrile participants was higher in Saraya than in Keur Socé (P < 0.001). It is also noteworthy that the use of insecticide-treated nets was significantly higher in Keur Socé than in Saraya (P < 0.001).
Table 2. Personal, Clinical and Epidemiological Characteristics of the Study Population.
| | Keur Soce | Saraya |
Sample (%) | | 545 | 561 |
Sex | F | 316 (57.98%) | 255 (45.45%) |
M | 229 (42.02%) | 306 (54.55%) |
Age (years) | Median ± IQR | 12 [10 - 25] | 13 [10 - 25] |
5 - 10 | 192 (35.23%) | 168 (29.95%) |
11 - 25 | 221 (40.55%) | 265 (47.24%) |
26 - 40 | 55 (10.09%) | 57 (10.16%) |
> 40 | 77 (14.13%) | 71 (12.65%) |
Axillary temperature C (Mean ±SD) | | 36.24 ± 0.61 | 36.52 ± 0.63 |
Fever (Body Temperature ≥ 37.5°C) | | 9 | 32 |
Hb (Mean ± SD) | | 12.17 ± 1.74 | 12.31 ± 1.73 |
Anaemia | | 19.82% | 15.51% |
Positive gametocytes (%) | | 0.18% | 1.24% |
Diagnostic | Plasmodium falciparum | 2.2% | 22.1% (124/561) |
Plasmodium malariae Plasmodium ovale | 0% 0% | 1.61% (9/561) 0.18% (1/561) |
PD (Median ± IQR) | | 1200 [715 - 2676] | 1105 [441.5 - 2229.5] |
RDT positive | | 2.75% (15/545) | 21.75% (122/561) |
Bed nets used | | 488 | 395 |
Beds nets used the last night | | 429 | 297 |
3.2. Parasitology
The study revealed more positive RDTs in Saraya than in Keur Socé, with prevalences of 21.75% and 2.75% respectively. Three species of Plasmodium were detected, with P. falciparum being the only one found in both localities; a prevalence of 2.2% in Keur Socé and 22.1% in Saraya. The other two species found in Saraya are P. malariae 1.61% and P. ovale 0.18%.
The prevalence of gametocytes observed by microscopy was very low in both Keur Socé (0.18%) and Saraya (1.24%).
The performance of the rapid diagnostic test (RDT) was assessed against microscopy, the established reference method. In Keur Soce, the RDT showed a sensitivity of 54.5% and a specificity of 98.3%. The positive predictive value (PPV) was 40.0%, while the negative predictive value (NPV) reached 99.1%. Overall accuracy was 97.4%.
In Saraya, sensitivity remained similar (56.7%), whereas specificity decreased to 88.5%. PPV reached 59.0% and NPV 87.5%, with an overall accuracy of 81.3%. These variations are consistent with previously reported site-specific differences in RDT performance.
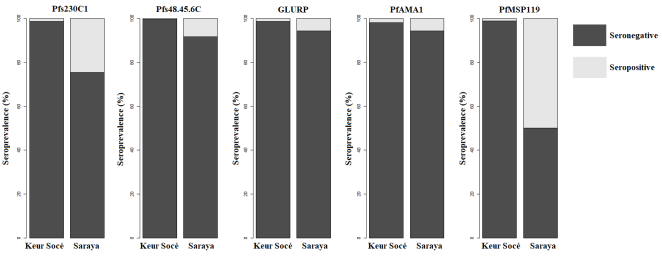
3.3. Antibody Response to P. falciparum Antigens in Keur Socé and Saraya
The seroprevalence of antibodies against
Plasmodium falciparum antigens differed markedly between Keur Socé and Saraya (
Table 3). For all antigens assessed, Saraya exhibited significantly higher serological responses (P ≤ 0.002).
For asexual blood-stage antigens, the seroprevalence of anti-PfAMA1 antibodies was 1.83% (10/545) in Keur Socé compared with 5.53% (31/561) in Saraya (P = 0.002). Similarly, anti-PfGLURP antibodies were detected in 1.28% (7/545) of participants in Keur Socé and 5.53% (31/561) in Saraya (P = 0.00021). An even larger difference was observed for PfMSP119, for which seroprevalence reached 1.1% (6/545) in Keur Socé versus 49.9% (280/561) in Saraya (P < 0.0001).
Marked contrasts were also observed for sexual-stage antigens. Anti-Pfs48/45-6C antibodies were detected in 0.37% (2/545) of individuals in Keur Socé compared with 8.2% (46/561) in Saraya (P < 0.0001). Likewise, anti-Pfs230C1 antibodies were present in 1.28% (7/545) of participants in Keur Socé and 24.6% (138/561) in Saraya (P < 0.0001).
Table 3. Seroprevalence of Antibodies to Plasmodium Falciparum Antigens.
Site | PfAMA1 | PfGLURP | Pfs48/45-6C | PfMSP119 | Pfs230C1 |
Keur Socé | 1.83% (10/545) | 1.28% (7/545) | 0.37% (2/545) | 1.1% (6/545) | 1.28% (7/545) |
Saraya | 5.53% (31/561) | 5.53% (31/561) | 8.2% (46/561) | 49.91% (280/561) | 24.6% (138/561) |
P | 0.002009 | 0.0002101 | <0.0001 | <0.0001 | <0.0001 |
The levels of specific IgG antibodies and seropositivity rates observed in Keur Socé and Saraya are presented in
Figures 1 and 2. Keur Socé is characterized by low seropositivity across all assessed antigens, whereas Saraya exhibits significantly higher rates, indicating greater exposure to
Plasmodium in this locality.
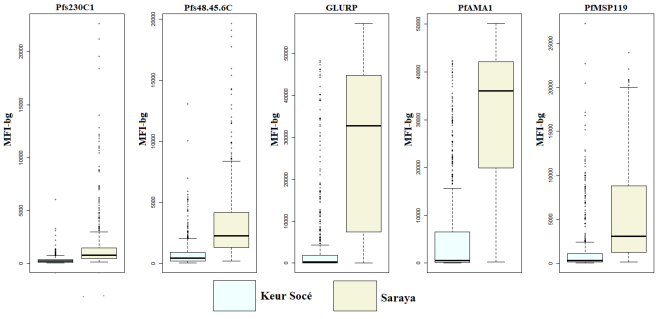
The median concentrations of all measured antibodies were significantly elevated in Saraya compared to Keur Socé. In Keur Socé, the highest median antibody responses were directed against PfAMA1, followed by Pfs48/45-6C. In Saraya, the strongest median responses were observed against PfAMA1 and PfGLURP (
Figure 2).
Figure 1. Seroprevalence of Antibodies to P. Falciparum Antigens in Keur Socé and Saraya.
Figure 2. Antibody Levels Against P. Falciparum Antigens in Keur Socé and Saraya.
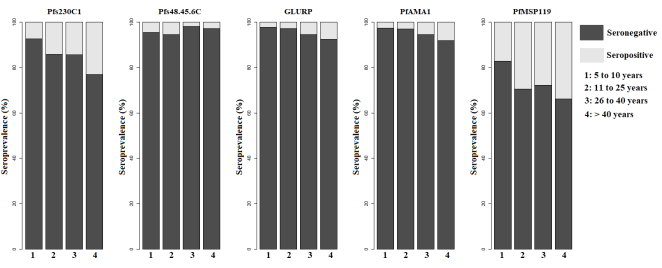
3.4. Effect of Age on Antibody Responses in Saraya
Given that antigen seroprevalence was extremely low in Keur Soce (between 0.37% and 1.83%) (
Table 3), it is difficult to draw meaningful conclusions from this site. In contrast, in Saraya, seroprevalence was significantly higher (P ≤ 0.002). Therefore, subsequent analyses of age-related correlations were restricted to this locality in order to evaluate the impact of age on naturally acquired immune responses (
Figure 3).
Seropositivity for anti-PfAMA1, anti-PfMSP1
119, anti-PfGLURP, and anti-Pfs230C1 antibodies increased considerably with age. Conversely, seropositivity for anti-Pfs48/45-6C antibodies was higher in the two youngest age groups (
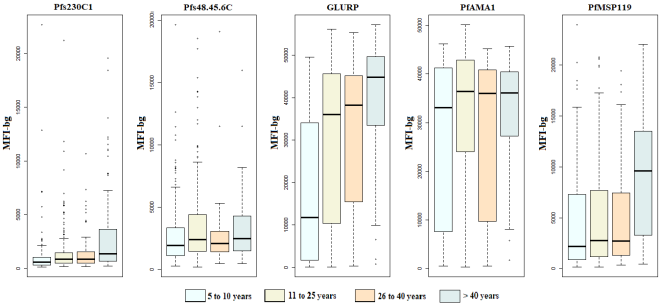
Figure 3). Median antibody levels against the three asexual-stage antigens also increased significantly with age, indicating cumulative exposure to Plasmodium infection. An age-dependent increase in antibody responses to the sexual-stage antigen Pfs230C1 was also observed; however, responses to Pfs48/45-6C did not differ significantly across age groups (
Figure 4).
Figure 3. Variation in Seropositive and Seronegative of Antibodies to P. Falciparum Antigens as a Function of Age in Saraya.
Figure 4. Variation in Antibody Concentrations Against P. Falciparum Antigens as a Function of Age in Saraya.
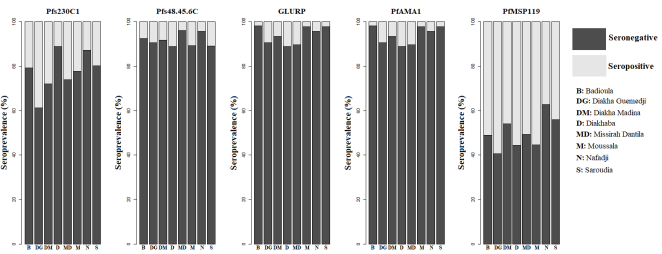
3.5. Antibody Responses Among Study Participants Across Saraya Villages
The seropositivity of anti-PfMSP1
19 antibodies was the highest across all villages (
Figure 5). It was followed by the seropositivity of the sexual-stage antibodies Pfs230C1 and Pfs48/45-6C. In contrast, the seropositivity of anti-GLURP and anti-PfAMA1 antibodies was very low in Badioula, Moussala, Nafadji, and Saroudia.
Figure 5. Variation in Seroprevalence of Antibodies to P. Falciparum Antigens in the Eight Villages of Saraya.
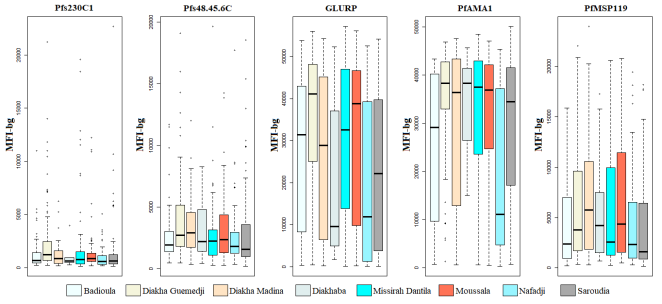
Except for anti-Pfs230C1 seroprevalance (P = 0.006), no statistical differences in seroprevalence between villages were observed. However, significant differences in antibody levels were observed between villages, and anti-PfAMA1 and anti-PfGLURP responses were greatest in all villages. Anti-Pfs48/45-6C levels were greater in magnitude than those of anti-Pfs230C1 all villages (
Figure 6).
Figure 6. Variation in Antibody Levels to P. Falciparum Antigens in the Eight Villages of Saraya.
3.6. Correlation of Antibody Responses Between the Different Antigens
The highest correlation (r > 0.7) between PfAMA1 and GLURP demonstrates that the same immune stimulus strongly stimulates these responses. The strong correlations between asexual antigens confirm the consistency of the immune response. The moderate associations between asexual and sexual antigens reflect the complexity of natural exposure. Overall, the high level of significance (p < 0.001) reinforces the reliability of the observations (
Table 4).
Table 4. Spearman’s Rank Correlation of Levels of Antibody Asexual Parasite Blood Stage Antigens with the Level of Antibody Sexual Parasite Stage Antigen.
| PfAMA1 | GLURP | Pfs48.45.6C | PfMSP119 | Pfs230C1 |
PfAMA1 | | 0.710*** | 0.411*** | 0.600*** | 0.321*** |
GLURP | | | 0.414*** | 0.610*** | 0.402*** |
Pfs48.45.6C | | | | 0.280*** | 0.360*** |
PfMSP119 | | | | | 0.300*** |
Pfs230C1 | | | | | |
3.7. Human- and Parasite-Related Factors, Associated with the Evolution of Immune Responses to Plasmodium spp.
Univariate analyses were performed to investigate the relationship between the individual variation in specific anti-
Plasmodium antibody responses and different demographic, biological and parasitological variables (
Table 5). Haemoglobin concentration, and parasite density on D were not associated with any variation in antibody response to
Plasmodium antigens (P > 0.05). Age was associated with variations in antibody against all antigens other than Pfs48.45.6C (P = 0.021).
Table 5. Influence of Covariates on the Responses of Antibody Levels to Various Plasmodium Antigens.
Covariate Factors | Antigens | R | P |
DP | Pfs230C1 | 0.003 | 0.976 |
| Pfs48.45.6C | -0.163 | 0.057 |
| PfGLURP | -0.108 | 0.206 |
| PfAMA1 | -0.080 | 0.351 |
| PfMSP119 | -0.079 | 0.357 |
Age | Pfs230C1 | 0.202 | <0.0001 |
| Pfs48.45.6C | 0.013 | 0.660 |
| PfGLURP | 0.206 | <0.0001 |
| PfAMA1 | 0.106 | 0.001 |
| PfMSP119 | 0.185 | <0.0001 |
Hb | Pfs230C1 | -0.018 | 0.547 |
| Pfs48.45.6C | -0.021 | 0.493 |
| PfGLURP | 0.020 | 0.186 |
| PfAMA1 | -0.034 | 0.262 |
| PfMSP119 | -0.029 | 0.331 |
4. Discussion
To improve our understanding of naturally acquired immunity after malaria exposure, we assessed the humoral response against P. falciparum antigens PfAMA1, PfMSP119, PfGLURP, Pfs230C1 and Pfs48.45.6C antigens of adults and children living in two malaria-endemic settings in Senegal with different levels of transmission. There were no differences in the age group and gender distributions of participants across the two sites. The most prevalent infecting plasmodial species was P. falciparum, reflecting the current malaria scenario in wider Senegal, where P. falciparum is responsible for most of infections.
Our study highlights contrasting malaria epidemiology between Keur Soce and Saraya. Saraya exhibited markedly higher
Plasmodium falciparum prevalence, fever, gametocyte carriage, and RDT positivity, confirming active transmission, consistent with findings from other high-transmission areas in West Africa
| [22] | Cissé B, Ba EH, Sokhna C, NDiaye JL, Gomis JF, Dial Y, et al. Effectiveness of Seasonal Malaria Chemoprevention in Children under Ten Years of Age in Senegal: A Stepped-Wedge Cluster-Randomised Trial. PLoS Med. nov 2016; 13(11): e1002175. |
[22]
. In contrast, Keur Soce showed low infection rates and higher anemia, suggesting lower and more heterogeneous transmission.
RDT performance varied by site: high specificity in Keur Soce minimized false positives, whereas reduced sensitivity in Saraya reflects the difficulty of detecting low-density infections in high-transmission areas, as reported elsewhere
| [23] | Wongsrichanalai, C., et al. (2007) A Review of Malaria Diagnostic Tools Microscopy and Rapid Diagnostic Test (RDT). The American Journal of Tropical Medicine and Hygiene, 77, 119-127. - References - Scientific Research Publishing [Internet]. [cité 30 nov 2025]. |
[23]
. The lower negative predictive value in Saraya indicates that RDTs alone may underestimate the infection burden, particularly asymptomatic carriers
| [24] | Bousema T, Okell L, Felger I, Drakeley C. Asymptomatic malaria infections: detectability, transmissibility and public health relevance. Nat Rev Microbiol. déc 2014; 12(12): 833-40. |
[24]
.
Despite similar anemia rates, parasite positivity was nearly tenfold higher in Saraya, suggesting frequent exposure rather than hematological vulnerability differences. Lower bed net use in Saraya likely contributes to higher transmission, supporting previous observations on the importance of preventive measures
| [25] | Lengeler C. Insecticide-treated bed nets and curtains for preventing malaria. Cochrane Database Syst Rev. 2004;(2): CD000363. |
[25]
.
These findings underscore malaria’s geographic heterogeneity and the need for locally tailored control strategies, combining RDTs, microscopy, and molecular tools for accurate epidemiological monitoring.
This study highlights marked differences in seroprevalence between Keur Socé and Saraya, reflecting highly contrasting levels of exposure to Plasmodium falciparum. The significantly higher seroprevalences observed in Saraya for all analyzed antigens indicate more sustained and active transmission in this area.
The asexual stage antigens PfAMA1 and PfGLURP, generally considered markers of relatively recent exposure, show values approximately three to four times higher in Saraya. This trend suggests more frequent circulation of clinical or subclinical infections in this locality. The extremely pronounced difference observed for PfMSP119, with seroprevalence reaching nearly 50% in Saraya compared to only 1.1% in Keur Socé, indicates much greater cumulative exposure. PfMSP119, known for eliciting stable, long-lasting antibody responses, often reflects a prolonged exposure history, reinforcing the hypothesis of persistent transmission in Saraya. Our results are consistent with the observations of
| [26] | McCallum FJ, Persson KEM, Fowkes FJI, Reiling L, Mugyenyi CK, Richards JS, et al. Differing rates of antibody acquisition to merozoite antigens in malaria: implications for immunity and surveillance. J Leukoc Biol. avr 2017; 101(4): 913-25. |
[26]
, who showed that, in general, antibody levels increase with transmission intensity.
The differences observed for the sexual stage antigens Pfs48/45-6C and Pfs230C1 also provide key insights into transmission dynamics. The high seroprevalences observed in Saraya (8.2% and 24.6%, respectively) suggest an increased presence of gametocytes and, consequently, a higher potential for vector transmission. In low-transmission contexts such as Keur Socé, the extremely low rates obtained (≤1.28%) confirm limited and potentially seasonal parasite circulation.
Regarding the evolution of the immune response with age, our analysis focused exclusively on antibody responses in the population of Saraya, an area with intense transmission. Except for the response against Pfs48.45.6C, age emerged as a significant predictor of antibody responses to PfAMA1, GLURP, PfMSP119, and Pfs230C1 (p < 0,001). Similar findings were reported in Tanzania by Bousema et al.
| [27] | Bousema T, Roeffen W, Meijerink H, Mwerinde H, Mwakalinga S, van Gemert GJ, et al. The dynamics of naturally acquired immune responses to Plasmodium falciparum sexual stage antigens Pfs230 & Pfs48/45 in a low endemic area in Tanzania. PloS One. 29 nov 2010; 5(11): e14114. |
[27]
, where a significant increase with age was observed for PfAMA1 (p < 0.001), PfMSP119 (p = 0.013), and Pfs230 (p = 0.006) after intra-individual adjustment.
In Ghana, a longitudinal study in children showed that anti-Pfs230 antibodies increased significantly with age: positive correlation (r=0.358; p=0.008). In children over 10 years of age, levels increased as early as March (p=0.002). Similar trends to the age effect were observed for Pfs48.45 (but with seasonal fluctuations and less age-related responses in certain phases)
| [28] | Broni FK, Acquah FK, Obiri-Yeboah D, Obboh EK, Sarpong E, Amoah LE. Profiling the Quality and Quantity of Naturally Induced Antibody Responses Against Pfs230 and Pfs48/45 Among Non-Febrile Children Living in Southern Ghana: A Longitudinal Study. Front Cell Infect Microbiol [Internet]. 25 nov 2021 [cité 17 oct 2025]; 11. |
[28]
.
In our study, the prevalence of anti-Pfs48.45.6C antibodies ranged from 0.37% to 8.2%, values far lower than those reported for the same antigen in central Ghana (74.7%)
| [29] | Acquah FK, Obboh EK, Asare K, Boampong JN, Nuvor SV, Singh SK, et al. Antibody responses to two new Lactococcus lactis-produced recombinant Pfs48/45 and Pfs230 proteins increase with age in malaria patients living in the Central Region of Ghana. Malar J. 1 août 2017; 16(1): 306. |
[29]
. These differences may reflect varying levels of exposure to gametocytes, as previously suggested in a longitudinal study conducted in a hypo-endemic area of Tanzania
| [27] | Bousema T, Roeffen W, Meijerink H, Mwerinde H, Mwakalinga S, van Gemert GJ, et al. The dynamics of naturally acquired immune responses to Plasmodium falciparum sexual stage antigens Pfs230 & Pfs48/45 in a low endemic area in Tanzania. PloS One. 29 nov 2010; 5(11): e14114. |
[27]
.
Age may be a modulating factor in the acquisition or maintenance of sexual stage immune responses after (recent) exposure to sexual stage antigens. The prevalence of antibodies against Pfs230C1 was higher in adults where they also appeared to have a longer half-life. More stable sexual stage immune responses in adults were previously reported in The Gambia
| [30] | Taylor RR, Egan A, McGuinness D, Jepson A, Adair R, Drakely C, et al. Selective recognition of malaria antigens by human serum antibodies is not genetically determined but demonstrates some features of clonal imprinting. Int Immunol. juin 1996; 8(6): 905-15. |
[30]
. There may be an innate difference between children and adults in how acquired immune systems function
| [31] | Yazdanbakhsh M, Sacks DL. Why does immunity to parasites take so long to develop? Nat Rev Immunol. févr 2010; 10(2): 80-1. |
[31]
, resulting in more efficient immune response after recent exposure in adults compared to children
| [32] | Giha HA, Nasr A, Iriemenam NC, Balogun HA, Arnot D, Theander TG, et al. Age-dependent association between IgG2 and IgG3 subclasses to Pf332-C231 antigen and protection from malaria, and induction of protective antibodies by sub-patent malaria infections, in Daraweesh. Vaccine. 17 févr 2010; 28(7): 1732-9. |
[32]
. In addition, higher cumulative exposure may have resulted in a more stable immune response in adults
| [30] | Taylor RR, Egan A, McGuinness D, Jepson A, Adair R, Drakely C, et al. Selective recognition of malaria antigens by human serum antibodies is not genetically determined but demonstrates some features of clonal imprinting. Int Immunol. juin 1996; 8(6): 905-15. |
[30]
. In the Saraya district, where seasonal malaria chemoprevention is implemented in children aged 3 months to 10 years
| [22] | Cissé B, Ba EH, Sokhna C, NDiaye JL, Gomis JF, Dial Y, et al. Effectiveness of Seasonal Malaria Chemoprevention in Children under Ten Years of Age in Senegal: A Stepped-Wedge Cluster-Randomised Trial. PLoS Med. nov 2016; 13(11): e1002175. |
[22]
, low antibody levels are observed. This may be explained by the prevention programme itself, as well as by the absence of repeated exposure to the parasite. Furthermore, studies conducted in Senegal have shown a reduction in the production of antimalarial antibodies in subjects who benefited from chemoprevention
| [17] | Sylla K, Tine RCK, Ndiaye M, Sow D, Sarr A, Mbuyi MLT, et al. Sero-epidemiological evaluation of Plasmodium falciparum malaria in Senegal. Malar J. 16 juill 2015; 14(1): 275. |
[17]
. Children who have exited the SMC programme need to be monitored, as they are at risk of developing severe malaria.
Across the villages of Saraya, no significant variation in the prevalence of anti-P. falciparum antibodies was observed, except for anti-Pfs230C1 antibodies. However, antibody levels differed substantially across antigens, with the highest responses recorded for PfAMA1 and PfGLURP. The antigen-specific nature of antibody responses
| [33] | Fowkes FJI, McGready R, Cross NJ, Hommel M, Simpson JA, Elliott SR, et al. New Insights into Acquisition, Boosting, and Longevity of Immunity to Malaria in Pregnant Women. J Infect Dis. 15 nov 2012; 206(10): 1612-21. |
[33]
may account for these differential dynamics. Parasite-, host-, and environment-related factors may also shape these responses and contribute to the differences observed between villages.
The significant variation in antibody levels among villages may be linked to the genetic background of the study populations, as allelic variation in major histocompatibility complex (HLA) antigens can influence the development of naturally acquired humoral immunity
| [34] | Lima-Junior J da C, Pratt-Riccio LR. Major Histocompatibility Complex and Malaria: Focus on Plasmodium vivax Infection. Front Immunol. 2016; 7: 13. |
| [35] | Pratt-Riccio LR, De Souza Perce-Da-Silva D, Da Costa Lima-Junior J, Pratt Riccio EK, Ribeiro-Alves M, Santos F, et al. Synthetic Antigens Derived from Plasmodium falciparum Sporozoite, Liver, and Blood Stages: Naturally Acquired Immune Response and Human Leukocyte Antigen Associations in Individuals Living in a Brazilian Endemic Area. Am J Trop Med Hyg. nov 2017; 97(5): 1581-92. |
[34, 35]
. Additionally, the genetic diversity of local P. falciparum isolates could also contribute, given that polymorphisms in B-cell epitopes may alter the immunological properties of antigens
| [36] | Pratt-Riccio LR, Perce-da-Silva D de S, Lima-Junior J da C, Theisen M, Santos F, Daniel-Ribeiro CT, et al. Genetic polymorphisms in the glutamate-rich protein of Plasmodium falciparum field isolates from a malaria-endemic area of Brazil. Mem Inst Oswaldo Cruz. juin 2013; 108(4): 523-8. |
[36]
. This hypothesis warrants further investigation.
The levels of antibodies against several plasmodial antigens seem to be associated with age to malaria in endemic regions
| [37] | Osier FHA, Fegan G, Polley SD, Murungi L, Verra F, Tetteh KKA, et al. Breadth and Magnitude of Antibody Responses to Multiple Plasmodium falciparum Merozoite Antigens Are Associated with Protection from Clinical Malaria. Infect Immun. mai 2008; 76(5): 2240-8. |
[37]
. In this study, individuals with higher levels of antibody were older and presented a longer time of residence in malaria-endemic areas than with levels of antibody of younger. Negative correlations were observed between parasitaemia and levels of antibody.
The limitations of our study included the low antibody levels obtained from Keur Socé, which prevented us from exploiting the data to examine characteristics related to our study population.
5. Conclusion
Our study demonstrated the prevalence of antibodies that recognized the five plasmodial antigens as well as the antibody levels increase with exposure to infection in two settings of high and low transmission settings in a malaria-endemic country. We also showed how these antibodies may contribute to parasite-induced natural immunity.
Children on SMC should also be monitored, as we have seen very low antibody secretion in this group. Additional studies are needed to confirm these findings.
Abbreviations
SMC | Seasonal Malaria Chemoprevention |
PfAMA1 | Plasmodium falciparum Apical Membrane Antigen 1 |
PfGLURP R2 | Plasmodium falciparum Glutamate-rich Protein Region 2 |
PfMSP119 | Plasmodium falciparum merozoite Surface Protein 1 (19-kDa carboxy-terminal fragment) |
RDT | Rapid Diagnostic Test |
CSP | Circumsporozoite Protein |
WBC | White Blood Cells |
DBS | Dried Blood Spot |
PBS | Phosphate Buffered Saline |
PBS/T | Phosphate Buffered Saline with Tween 20 |
MFI | Median Fluorescence Intensity |
MFI-bg | Median Fluorescence Intensity Background-corrected Data |
NPV | Negative Predictive Value |
PPV | Positive Predictive Value |
Acknowledgments
Work was funded by Malaria Research Capacity Development in West and Central Africa. The publication costs were covered by the parasitology and mycology department of Cheikh Anta Diop University of Dakar. We thank Matthijs Jore and Roos de Jong (Radboud UMC, Nijmegen, the Netherlands) for production of 48/45-6C and Jessica Chichester (Fraunhofer) Pfs230CMB.
Author Contributions
Lam Aminata: Data curation, Formal Analysis, Funding acquisition, Methodology, Writing – original draft, Writing – review & editing
Lo Aminata Colle: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Methodology, Supervision, Validation, Writing – review & editing
Patterson Catriona: Data curation, Formal Analysis, Methodology, Writing – review & editing
Sylla Khadime: Data curation, Formal Analysis, Writing – review & editing
Kebe Khadim: Data curation, Formal Analysis, Writing – review & editing
Gaye Ndeye Aida: Methodology, Writing – review & editing
Manga Isaac Akhenaton: Methodology, Writing – review & editing
Lelo Souleye: Methodology, Writing – review & editing
Fall Cheikh Binetou: Methodology, Writing – review & editing
Diouf Marie Pierre: Methodology, Writing – review & editing
Minlekib Carole Pab: Methodology, Writing – review & editing
Tine Roger Clement: Conceptualization, Methodology, Writing – review & editing
Drakeley Chris: Conceptualization, Data curation, Formal Analysis, Methodology, Supervision, Validation, Writing – review & editing
Faye Babacar: Conceptualization, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing
Conflicts of Interest
The authors declare that they have no competing interest.
References
| [1] |
Combemorel P. Le cycle de vie de Plasmodium falciparum | Planet-Vie [The life cycle of Plasmodium falciparum | Planet-Life]. 2019 [cité 30 nov 2025].
|
| [2] |
Barry A, Hansen D. Naturally acquired immunity to malaria. Parasitology. févr 2016; 143(2): 125-8.
|
| [3] |
Epidemiology and Infectivity of Plasmodium falciparum and Plasmodium vivax Gametocytes in Relation to Malaria Control and Elimination | Clinical Microbiology Reviews [Internet]. [cité 30 nov 2025].
|
| [4] |
Khan SM, Franke-Fayard B, Mair GR, Lasonder E, Janse CJ, Mann M, et al. Proteome analysis of separated male and female gametocytes reveals novel sex-specific Plasmodium biology. Cell. 3 juin 2005; 121(5): 675-87.
|
| [5] |
van Dijk MR, Janse CJ, Thompson J, Waters AP, Braks JA, Dodemont HJ, et al. A central role for P48/45 in malaria parasite male gamete fertility. Cell. 12 janv 2001; 104(1): 153-64.
|
| [6] |
Tachibana M, Miura K, Takashima E, Morita M, Nagaoka H, Zhou L, et al. Identification of domains within Pfs230 that elicit transmission blocking antibody responses. Vaccine. 22 mars 2019; 37(13): 1799-806.
|
| [7] |
Sauerwein RW, Bousema T. Transmission blocking malaria vaccines: Assays and candidates in clinical development. Vaccine. 22 déc 2015; 33(52): 7476-82.
|
| [8] |
Yazdani SS, Mukherjee P, Chauhan VS, Chitnis CE. Immune responses to asexual blood-stages of malaria parasites. Curr Mol Med. mars 2006; 6(2): 187-203.
|
| [9] |
Healer J, Chiu CY, Hansen DS. Mechanisms of naturally acquired immunity to P. falciparum and approaches to identify merozoite antigen targets. Parasitology. juin 2018; 145(7): 839-47.
|
| [10] |
De Jong RM, Tebeje SK, Meerstein-Kessel L, Tadesse FG, Jore MM, Stone W, et al. Immunity against sexual stage Plasmodium falciparum and Plasmodium vivax parasites. Immunol Rev. janv 2020; 293(1): 190-215.
|
| [11] |
Ouédraogo AL, Roeffen W, Luty AJF, de Vlas SJ, Nebie I, Ilboudo-Sanogo E, et al. Naturally Acquired Immune Responses to Plasmodium falciparum Sexual Stage Antigens Pfs48/45 and Pfs230 in an Area of Seasonal Transmission ▿. Infect Immun. déc 2011; 79(12): 4957-64.
|
| [12] |
Kengne-Ouafo JA, Sutherland CJ, Binka FN, Awandare GA, Urban BC, Dinko B. Immune Responses to the Sexual Stages of Plasmodium falciparum Parasites. Front Immunol [Internet]. 1 févr 2019 [cité 30 nov 2025]; 10.
|
| [13] |
Linn AM, Ndiaye Y, Hennessee I, Gaye S, Linn P, Nordstrom K, et al. Reduction in symptomatic malaria prevalence through proactive community treatment in rural Senegal. Trop Med Int Health TM IH. nov 2015; 20(11): 1438-46.
|
| [14] |
Ndiaye Y, Ndiaye JLA, Cisse B, Blanas D, Bassene J, Manga IA, et al. Community case management in malaria: review and perspectives after four years of operational experience in Saraya district, south-east Senegal. Malar J. 12 juill 2013; 12: 240.
|
| [15] |
Agence Nationale de la Statistique et de la Démographie (ANSD) et ICF International. Enquête Démographique et de Santé à Indicateurs Multiples Sénégal (EDS-MICS) 2010-2011 [National Agency for Statistics and Demography (ANSD) and ICF International. Senegal Multiple Indicator Cluster Survey (MICS-EDS) 2010-2011]. Calverton, Maryland, USA; 2012. (ANSD et ICF International).
|
| [16] |
Ndiath M, Faye B, Cisse B, Ndiaye JL, Gomis JF, Dia AT, et al. Identifying malaria hotspots in Keur Soce health and demographic surveillance site in context of low transmission. Malar J. 24 nov 2014; 13(1): 453.
|
| [17] |
Sylla K, Tine RCK, Ndiaye M, Sow D, Sarr A, Mbuyi MLT, et al. Sero-epidemiological evaluation of Plasmodium falciparum malaria in Senegal. Malar J. 16 juill 2015; 14(1): 275.
|
| [18] |
World Health Organization. World malaria report 2020 [Internet]. [cité 17 oct 2025]. Report No.: 978-92-4-001579-1.
|
| [19] |
Muerhoff AS, Birkenmeyer LG, Coffey R, Dille BJ, Barnwell JW, Collins WE, et al. Detection of Plasmodium falciparum, P. vivax, P. ovale, and P. malariae Merozoite Surface Protein 1-p19 Antibodies in Human Malaria Patients and Experimentally Infected Nonhuman Primates. Clin Vaccine Immunol CVI. oct 2010; 17(10): 1631-8.
|
| [20] |
Rogier E, van den Hoogen L, Herman C, Gurrala K, Joseph V, Stresman G, et al. High-throughput malaria serosurveillance using a one-step multiplex bead assay. Malar J. 4 déc 2019; 18(1): 402.
|
| [21] |
Eacret JS, Parzych EM, Gonzales DM, Burns JM. Inclusion of an optimized Plasmodium falciparum merozoite surface protein 2-based antigen in a trivalent, multi-stage malaria vaccine. J Immunol Baltim Md 1950. 15 avr 2021; 206(8): 1817-31.
|
| [22] |
Cissé B, Ba EH, Sokhna C, NDiaye JL, Gomis JF, Dial Y, et al. Effectiveness of Seasonal Malaria Chemoprevention in Children under Ten Years of Age in Senegal: A Stepped-Wedge Cluster-Randomised Trial. PLoS Med. nov 2016; 13(11): e1002175.
|
| [23] |
Wongsrichanalai, C., et al. (2007) A Review of Malaria Diagnostic Tools Microscopy and Rapid Diagnostic Test (RDT). The American Journal of Tropical Medicine and Hygiene, 77, 119-127. - References - Scientific Research Publishing [Internet]. [cité 30 nov 2025].
|
| [24] |
Bousema T, Okell L, Felger I, Drakeley C. Asymptomatic malaria infections: detectability, transmissibility and public health relevance. Nat Rev Microbiol. déc 2014; 12(12): 833-40.
|
| [25] |
Lengeler C. Insecticide-treated bed nets and curtains for preventing malaria. Cochrane Database Syst Rev. 2004;(2): CD000363.
|
| [26] |
McCallum FJ, Persson KEM, Fowkes FJI, Reiling L, Mugyenyi CK, Richards JS, et al. Differing rates of antibody acquisition to merozoite antigens in malaria: implications for immunity and surveillance. J Leukoc Biol. avr 2017; 101(4): 913-25.
|
| [27] |
Bousema T, Roeffen W, Meijerink H, Mwerinde H, Mwakalinga S, van Gemert GJ, et al. The dynamics of naturally acquired immune responses to Plasmodium falciparum sexual stage antigens Pfs230 & Pfs48/45 in a low endemic area in Tanzania. PloS One. 29 nov 2010; 5(11): e14114.
|
| [28] |
Broni FK, Acquah FK, Obiri-Yeboah D, Obboh EK, Sarpong E, Amoah LE. Profiling the Quality and Quantity of Naturally Induced Antibody Responses Against Pfs230 and Pfs48/45 Among Non-Febrile Children Living in Southern Ghana: A Longitudinal Study. Front Cell Infect Microbiol [Internet]. 25 nov 2021 [cité 17 oct 2025]; 11.
|
| [29] |
Acquah FK, Obboh EK, Asare K, Boampong JN, Nuvor SV, Singh SK, et al. Antibody responses to two new Lactococcus lactis-produced recombinant Pfs48/45 and Pfs230 proteins increase with age in malaria patients living in the Central Region of Ghana. Malar J. 1 août 2017; 16(1): 306.
|
| [30] |
Taylor RR, Egan A, McGuinness D, Jepson A, Adair R, Drakely C, et al. Selective recognition of malaria antigens by human serum antibodies is not genetically determined but demonstrates some features of clonal imprinting. Int Immunol. juin 1996; 8(6): 905-15.
|
| [31] |
Yazdanbakhsh M, Sacks DL. Why does immunity to parasites take so long to develop? Nat Rev Immunol. févr 2010; 10(2): 80-1.
|
| [32] |
Giha HA, Nasr A, Iriemenam NC, Balogun HA, Arnot D, Theander TG, et al. Age-dependent association between IgG2 and IgG3 subclasses to Pf332-C231 antigen and protection from malaria, and induction of protective antibodies by sub-patent malaria infections, in Daraweesh. Vaccine. 17 févr 2010; 28(7): 1732-9.
|
| [33] |
Fowkes FJI, McGready R, Cross NJ, Hommel M, Simpson JA, Elliott SR, et al. New Insights into Acquisition, Boosting, and Longevity of Immunity to Malaria in Pregnant Women. J Infect Dis. 15 nov 2012; 206(10): 1612-21.
|
| [34] |
Lima-Junior J da C, Pratt-Riccio LR. Major Histocompatibility Complex and Malaria: Focus on Plasmodium vivax Infection. Front Immunol. 2016; 7: 13.
|
| [35] |
Pratt-Riccio LR, De Souza Perce-Da-Silva D, Da Costa Lima-Junior J, Pratt Riccio EK, Ribeiro-Alves M, Santos F, et al. Synthetic Antigens Derived from Plasmodium falciparum Sporozoite, Liver, and Blood Stages: Naturally Acquired Immune Response and Human Leukocyte Antigen Associations in Individuals Living in a Brazilian Endemic Area. Am J Trop Med Hyg. nov 2017; 97(5): 1581-92.
|
| [36] |
Pratt-Riccio LR, Perce-da-Silva D de S, Lima-Junior J da C, Theisen M, Santos F, Daniel-Ribeiro CT, et al. Genetic polymorphisms in the glutamate-rich protein of Plasmodium falciparum field isolates from a malaria-endemic area of Brazil. Mem Inst Oswaldo Cruz. juin 2013; 108(4): 523-8.
|
| [37] |
Osier FHA, Fegan G, Polley SD, Murungi L, Verra F, Tetteh KKA, et al. Breadth and Magnitude of Antibody Responses to Multiple Plasmodium falciparum Merozoite Antigens Are Associated with Protection from Clinical Malaria. Infect Immun. mai 2008; 76(5): 2240-8.
|
Cite This Article
-
APA Style
Aminata, L., Colle, L. A., Catriona, P., Khadime, S., Khadime, K., et al. (2025). Naturally Acquired Immune Responses to Antigens of the Sexual and Asexual Stages of Plasmodium Falciparum in the Southern and Central Regions of Senegal. International Journal of Infectious Diseases and Therapy, 10(4), 105-117. https://doi.org/10.11648/j.ijidt.20251004.14
 Copy
|
Copy
|
 Download
Download
ACS Style
Aminata, L.; Colle, L. A.; Catriona, P.; Khadime, S.; Khadime, K., et al. Naturally Acquired Immune Responses to Antigens of the Sexual and Asexual Stages of Plasmodium Falciparum in the Southern and Central Regions of Senegal. Int. J. Infect. Dis. Ther. 2025, 10(4), 105-117. doi: 10.11648/j.ijidt.20251004.14
Copy
|
Download
AMA Style
Aminata L, Colle LA, Catriona P, Khadime S, Khadime K, et al. Naturally Acquired Immune Responses to Antigens of the Sexual and Asexual Stages of Plasmodium Falciparum in the Southern and Central Regions of Senegal. Int J Infect Dis Ther. 2025;10(4):105-117. doi: 10.11648/j.ijidt.20251004.14
Copy
|
Download
-
@article{10.11648/j.ijidt.20251004.14,
author = {Lam Aminata and Lo Aminata Colle and Patterson Catriona and Sylla Khadime and Kebe Khadime and Gaye Ndeye Aida and Manga Isaac Akhenaton and Lelo Souleye and Fall Cheikh Binetou and Diouf Marie Pierre and Minlekib Carole Pab and Tine Roger Clement and Gaye Omar and Drakeley Chris and Faye Babacar},
title = {Naturally Acquired Immune Responses to Antigens of the Sexual and Asexual Stages of Plasmodium Falciparum in the Southern and Central Regions of Senegal},
journal = {International Journal of Infectious Diseases and Therapy},
volume = {10},
number = {4},
pages = {105-117},
doi = {10.11648/j.ijidt.20251004.14},
url = {https://doi.org/10.11648/j.ijidt.20251004.14},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijidt.20251004.14},
abstract = {In Senegal, despite numerous malaria control interventions, transmission is still seasonal. Malaria transmission depends on the presence of infectious parasites in the sexual stage in human peripheral blood. Immune responses acquired naturally at these or other stages can affect malaria transmission, resulting in protection against malaria, reduced transmission, and also form the basis for the development of transmission-blocking vaccines. To evaluate the antibody response profile against the asexual antigens PfAMA1, PfMSP119, Pf GLURP R2 and the sexual antigens Pfs230C1, Pfs48.45.6C in inhabitants naturally exposed to malaria in areas with different levels of transmission in Senegal. A cross-sectional study was carried out at the end of the transmission season in central (Keur Socé) and southern (Saraya) Senegal in 2018. We included 1106 asymptomatic volunteers aged 5 and over. Capillary blood was collected from each participant for an RDT, 2 slides for microscopy and a dried blood spot samples for immunology. A Luminex serological multiplex bead assay was then used to assess Plasmodium falciparum seroprevalence Our study population was characterized by a very young population with a median age of 12 15 years. The parasite prevalence of Plasmodium falciparum was 21.75% and 2.75% by RDT and 22.1% and 2.2% by microscopy for the southern and central regions respectively. Two other plasmodial species were found in Saraya, with prevalences of 1.61% for P. malariae and 0.18% for P. ovale. The mean seroprevalences of antibodies against three asexual blood-stage antigens (PfAMA1, PfGLURP and PfMSP119) and two sexualstage antigens (Pfs48.45.6C and Pfs230C1) were significantly higher in Saraya. In Keur Socé, the mean seroprevalence of antibodies against the PfAMA1 antigen was highest (1.83%), while in Saraya, PfMSP119 was highest (49.91%). The antigenicity of these proteins depended on endemicity levels, as antibody prevalence was statistically different in the two sites and increased with transmission intensity. With the exception of anti-Pfs48.45.6C antibody levels, all other antibody responses increased with age. Overall, these data indicate that the seroprevalence and antivody levels of individuals with antibodies recognizing all five antigens increase with exposure to infection, and that these antibodies may contribute to immunity against parasites. Children receiving SMC should also be monitored, as we have noted a loss of immunity in this group.},
year = {2025}
}
Copy
|
Download
-
TY - JOUR
T1 - Naturally Acquired Immune Responses to Antigens of the Sexual and Asexual Stages of Plasmodium Falciparum in the Southern and Central Regions of Senegal
AU - Lam Aminata
AU - Lo Aminata Colle
AU - Patterson Catriona
AU - Sylla Khadime
AU - Kebe Khadime
AU - Gaye Ndeye Aida
AU - Manga Isaac Akhenaton
AU - Lelo Souleye
AU - Fall Cheikh Binetou
AU - Diouf Marie Pierre
AU - Minlekib Carole Pab
AU - Tine Roger Clement
AU - Gaye Omar
AU - Drakeley Chris
AU - Faye Babacar
Y1 - 2025/12/30
PY - 2025
N1 - https://doi.org/10.11648/j.ijidt.20251004.14
DO - 10.11648/j.ijidt.20251004.14
T2 - International Journal of Infectious Diseases and Therapy
JF - International Journal of Infectious Diseases and Therapy
JO - International Journal of Infectious Diseases and Therapy
SP - 105
EP - 117
PB - Science Publishing Group
SN - 2578-966X
UR - https://doi.org/10.11648/j.ijidt.20251004.14
AB - In Senegal, despite numerous malaria control interventions, transmission is still seasonal. Malaria transmission depends on the presence of infectious parasites in the sexual stage in human peripheral blood. Immune responses acquired naturally at these or other stages can affect malaria transmission, resulting in protection against malaria, reduced transmission, and also form the basis for the development of transmission-blocking vaccines. To evaluate the antibody response profile against the asexual antigens PfAMA1, PfMSP119, Pf GLURP R2 and the sexual antigens Pfs230C1, Pfs48.45.6C in inhabitants naturally exposed to malaria in areas with different levels of transmission in Senegal. A cross-sectional study was carried out at the end of the transmission season in central (Keur Socé) and southern (Saraya) Senegal in 2018. We included 1106 asymptomatic volunteers aged 5 and over. Capillary blood was collected from each participant for an RDT, 2 slides for microscopy and a dried blood spot samples for immunology. A Luminex serological multiplex bead assay was then used to assess Plasmodium falciparum seroprevalence Our study population was characterized by a very young population with a median age of 12 15 years. The parasite prevalence of Plasmodium falciparum was 21.75% and 2.75% by RDT and 22.1% and 2.2% by microscopy for the southern and central regions respectively. Two other plasmodial species were found in Saraya, with prevalences of 1.61% for P. malariae and 0.18% for P. ovale. The mean seroprevalences of antibodies against three asexual blood-stage antigens (PfAMA1, PfGLURP and PfMSP119) and two sexualstage antigens (Pfs48.45.6C and Pfs230C1) were significantly higher in Saraya. In Keur Socé, the mean seroprevalence of antibodies against the PfAMA1 antigen was highest (1.83%), while in Saraya, PfMSP119 was highest (49.91%). The antigenicity of these proteins depended on endemicity levels, as antibody prevalence was statistically different in the two sites and increased with transmission intensity. With the exception of anti-Pfs48.45.6C antibody levels, all other antibody responses increased with age. Overall, these data indicate that the seroprevalence and antivody levels of individuals with antibodies recognizing all five antigens increase with exposure to infection, and that these antibodies may contribute to immunity against parasites. Children receiving SMC should also be monitored, as we have noted a loss of immunity in this group.
VL - 10
IS - 4
ER -
Copy
|
Download